
高等植物 RNA サイレンシングの研究
RNA サイレンシングの基本メカニズム
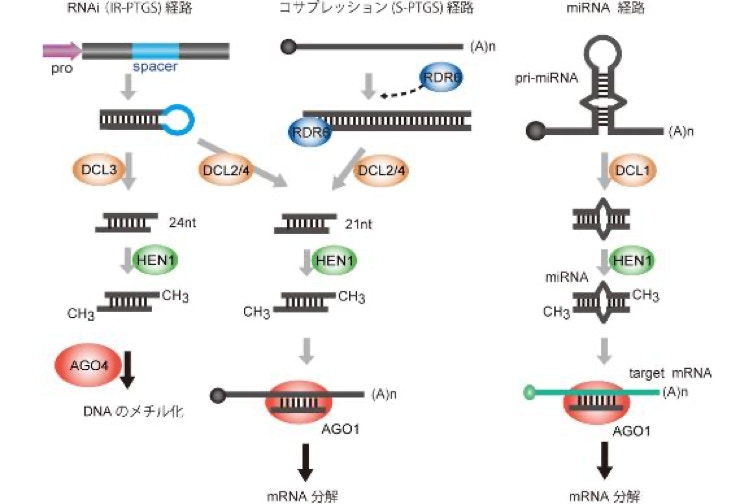
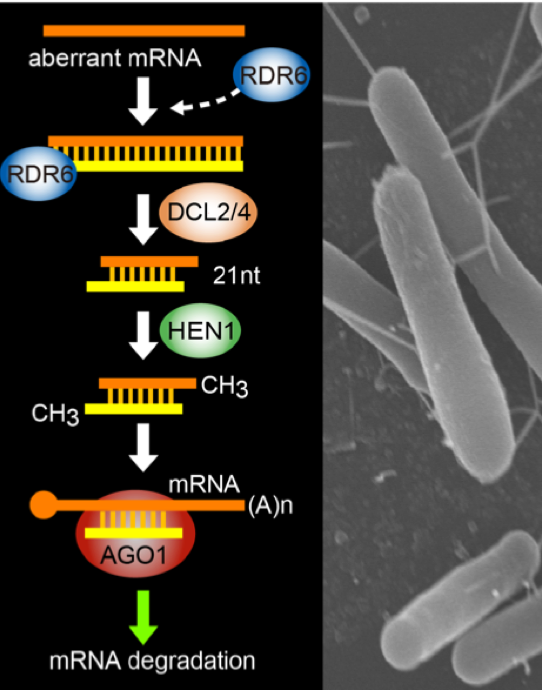
RNAサイレンシングと呼ばれるこの現象とその類似の現象は植物にとどまらず幅広い範囲の生物で観察され、世界 中で活発に研究がなされています。鍵を握る分子は21塩基から24塩基程度の短い鎖長からなるRNA分子(短鎖RNA)です。RNAサイレンシングでは、転写後RNAが短鎖RNA分子によるガイドを受けて分解されたり、短鎖RNAがDNAのメチル化などを誘導してクロマチン構造が変化します。短鎖RNAが本来ゲノムにコードされていない場合、その短鎖RNAはsiRNAと呼ばれます。一方、短鎖RNA自体がゲノムにコードされている場合、miRNAと呼ばれます。miRNAは発生や分化に重要な役割を果たしています。
このRNAサイレンシングを人為的にコントロールする方法が、RNA干渉(RNAi)です。RNA干渉は、Dicer-RISCシステムによる塩基配列特異的なRNA分解系です。 ファイアー・メロー両博士は1998年に線虫において、センスRNAとアンチセンスRNAを両方同時に投与すると、用いた配列の遺伝子の発現が抑制される ことを見出しました。彼らはこの現象にRNA干渉と名前を付け、現在では真核生物に共通して存在する遺伝子発現調節機構であることが明らかになっていま す。両博士は2006年ノーベル医学生理学賞を受賞されました。
S-PTG

研究室では、α-リノレン酸合成を触媒するω-3脂肪酸不飽和化酵素遺伝子を導入したタバコ形質転換株を作成しました。すると、ほとんどのタバコ株はα-リノレン酸含量が増加しました。
しかし、一部にα-リノレン酸含量が野生株よりも大幅に減少した株が得られました。これは、RNAサイレンシングにより、ω-3脂肪酸不飽和化酵素遺伝子に対するsiRNAが形成されたことが原因です。
(Tomita et al. (2004) FEBS Lett. 573: 117-120)
この現象は、コサプレッションもしくはS-PTGS (sense-transgene induced post-transcriptional gene silencing)と呼ばれます。
S-PTGに見られる器官特異性とは?
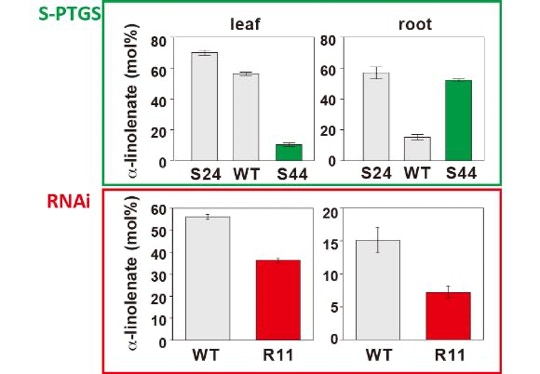
S-PTGSには、まだよくわかっていないことが多くあります。右の図は、ω-3脂肪酸不飽和化酵素遺伝子を標的としたS-PTGSと、人為的にRNA干渉を誘導したRNAi株をタバコで作成した結果です。S44株は、S-PTGS。RNAi株はR11株です。どちらのケースでもsiRNAは葉と根の両方で作られていることが確認されています。しかし、S-PTGS株であるS44株では、根ではα-リノレン酸含量が大幅に増加していることがわかります。その増加の程度は、過剰発現株であるS24株に匹敵します。一方、RNAi株であるR11株では、葉でも根でもα-リノレン酸含量は低くなりました。このことは、S-PTGSには、器官特異的な制御機構があることを示しています。でもその仕組みはまだまったくわかっていません。
研究室では、この器官特異的サイレンシングの仕組み、サイレンシング抑制因子、また翻訳段階でのサイレンシングなどの研究を進めています。