<略歴>
岩手県遠野市出身/岩手大学大学院農学研究科修士課程修了/名古屋大学大学院農学研究科博士課程満了/ワシントン大学生物学科(アメリカ合衆国ミズーリ州)/(株)植物工学研究所・博士研究員/国立遺伝学研究所・助手となる。次いで、(財)岩手生物工学研究センター・主席研究員を経て、1998年千葉大学大学院自然科学研究科の助教授として赴任、2009年4月より園芸学研究科の准教授に配置換え。2014年4月より教授。
<研究経緯>
専門は植物育種学、植物分子遺伝学、植物分子分類学。
学生時代から植物の進化に興味を持ち、博士論文のテーマは、「ダイズの起源」、その後、生活のために渡米して、遺伝子組み換えの仕事に従事する。それ以来、「植物の種分化」および「遺伝子組換えによる園芸植物の改良」に関する2つの研究に取り組んでいる。
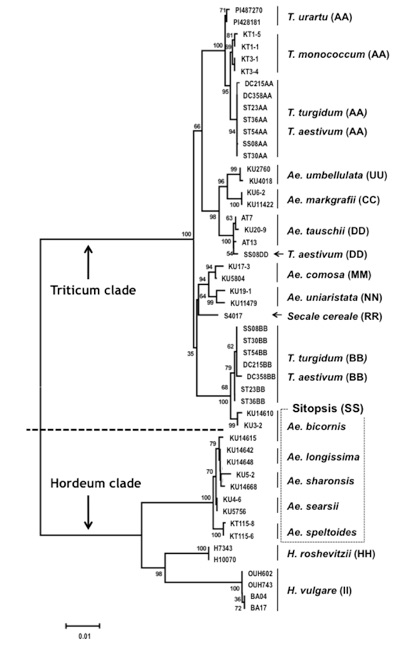
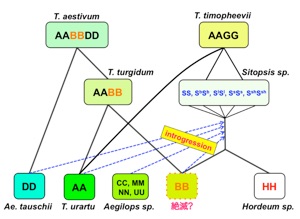
私の種分化に関する研究の特徴は、特定の植物に固有の現象を解析するのではなく、すべての植物に共通して適応できる技術を開発することである。PCR法を用いたDNAフィンガープリント法(ALPHA法1990)、植物の葉緑体の進化を解析するためのPSID (plastid subtype identity) 配列 (1997)、新たなゲノムウォーキング法 (Straight walk 2005)、真核生物の種に特異的なアミノ酸配列の解析法 (2010)などである。これらの手法を駆使し、園芸植物の種分化を解明することを目標としている。
また、アメリカ留学中に習得した遺伝子組換え技術を用いた研究もおこなっている。遺伝子組換え技術は、植物の遺伝子の機能を解明するための基礎実験に必要であるが、ストレス抵抗性など有用な形質を付与した園芸植物を作出するためにも有効である。また、植物のバイオマスを増大させるための遺伝子組換え技術の開発も試みている。