准教授 井川 智子(Tomoko Igawa)
メール:tigawa(*)chiba-u.jp
メールは (*)の部分を@マークにして下さい。
准教授 井川 智子(Tomoko Igawa)
メール:tigawa(*)chiba-u.jp
メールは (*)の部分を@マークにして下さい。
<略歴>
日本女子大学理学部卒業。千葉大学大学院自然科学研究科博士課程にて学位を取得。 その後理化学研究所、奈良先端科学技術大学院大学などの研究員を経て、2011年4月千葉大学大学院園芸学研究科に着任。


<研究経緯>
日本女子大学理学部物質生物科学科在学中に、故・駒嶺穆先生の講義で‘植物の分化全能性’を知り、植物のダイナミックな能力に感嘆しました。この分化全能性を利用してバイオテクノロジー育種に挑戦している三位正洋先生の研究に強く惹かれ、千葉大学大学院自然科学研究科(現在は園芸学研究科)に進学しました。博士課程ではスターチスという花き園芸植物を材料に、培養細胞を介した植物体再生の技術と遺伝子導入技術の開発研究で学位を取得しました。卒業後は穀類の組換え植物開発研究にも携わりました。
遺伝子組換え技術は、交雑による育種が困難な場合に直接遺伝子を導入して形質を変化させる、という点で非常に画期的な技術です。しかし実際には植物体再生も遺伝子導入も困難な植物種が多いのが現実です(比較的簡単にうまくいくのが‘モデル植物’と呼ばれます)。様々な植物種において遺伝子組換え技術を開発していく研究は実用化のためだけでなく、遺伝子機能を解析する基礎研究の促進のためにも重要であり、当研究室でも挑戦し続けています。
博士課程時代の材料、スターチス。
(Limonium sinuatum ‘Early Rose’)
培養には結構苦労しました。
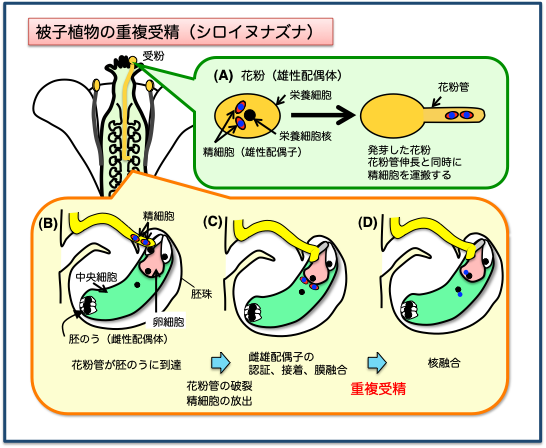
古典的な植物育種法の一つである掛け合わせは、植物の有性生殖現象を利用してきたものです。特に被子植物は、重複受精と呼ばれる独特の受精様式を進化させています。しかし意外にも重複受精の仕組みは、まだまだ謎の部分が多いのです。改めて(遠い将来ではありますが)育種への応用に繋がることを目指して、この現象を解明する研究を始めたところです。

テッポウユリの花粉管発芽
陸上植物の7割以上を占める被子植物は、私たちの生活にも必須の生物種です。被子植物は重複受精によって種子を作り、次世代を残します。
花粉は雌ずいの柱頭に付くと(受粉)、花粉管を発芽します。花粉管は伸長しながら雌ずい中を進み、個々の胚珠を目指します。花粉管はさらに胚珠の中の胚のう(雌性配偶体)に進入します。そこで花粉管が破裂して、運んできた2つの精細胞(雄性配偶子)を胚のう内に放出します。その後、2つの精細胞はそれぞれ卵細胞または中央細胞(雌性配偶子)と融合して、受精が行われます。このように、2種類の配偶子ペアの受精がほぼ同時に起こることから‘重複受精’と呼ばれています。
重複受精は1898年にロシアのNawaschinが発見しました。発見から100年以上経った今も、2つの精細胞がどのように卵細胞と中央細胞を「見分け」て、どのようにそれぞれとの受精に至るのか、その現象を支配する仕組みはほとんど明らかになっていません。
モデル植物であるシロイヌナズナは胚珠が小さいため、顕微鏡で観察しやすい、という利点があります。配偶子特異的に蛍光タンパク質を発現させて受精中の配偶子の構造変化を直接観察することで、重複受精の仕組みを細胞学的に解析する研究を行っています。さらに重複受精を制御する遺伝子を探索し、分子生物学的な解析も行っています。
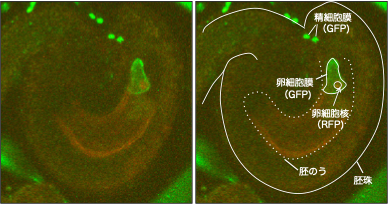
精細胞膜、卵細胞膜を緑色蛍光タンパク質(GFP)で、卵細胞核を赤色蛍光タンパク質RFPで可視化。
花粉管が胚珠に到達して胚のう内に近づいている段階の様子。
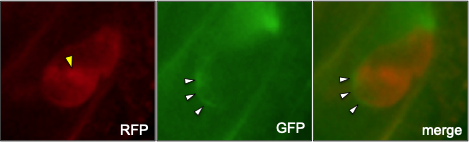
受精後の卵細胞(卵細胞膜をRFPで可視化)。精細胞核を示すRFPシグナル(黄色矢頭)は卵細胞の中に進入しているが、精細胞膜を示すGFPシグナル(白矢頭)は卵細胞の表面に残っている様子が観察される。雌雄配偶子の細胞膜融合が起きたことを示す(Igawa et al., 2013)。

研究内容:重複受精制御機構の解明









千葉大学ファイトケミカル植物分子科学に参画しています。
↑ロゴマークをクリックするとリンク先へ移動します↑
2017年度より新学術領域「植物新種誕生の原理」に参加しています。


2018年に新しい受精因子を同定しました!
千葉大学ニュースリリース
←画像をクリックするとリンクします
関連コラムをこちらに書いています
被子植物が2組のカップルを正確につくるための、男女のやりとり – 「重複受精」の新たな因子を探る
-アカデミストジャーナル(画像クリックでリンク→)
